转自:中国科学院海洋生物标本馆
原创:李宇航
链接:硅藻——海洋中的生命奇迹
“Few objects are more beautiful than the minute siliceous cases of diatoms: were they only created to be admired under the microscope?”很少有事物可以比硅藻精细的硅质外壳更加美丽:它们难道只是为了在显微镜下供人欣赏而生的吗?[Darwin, Charles. On the Origin of Species by Means of NaturalSelection. J. Murray, London, 1859]查尔斯·达尔文:物种起源
硅藻是多样性最高的单细胞真核藻类,现存超过250属,估计有10万余种。硅藻细胞大小跨度极大,从小于1微米至几毫米不等。硅藻分布于几乎所有水体环境中,从热带到极地,从强酸性到强碱性,从淡水到盐湖,从两极到温泉,甚至在土壤、树木的表面都有硅藻的踪迹。硅藻最显著的特征是其多样的硅质外壳(frustule)。硅藻的外壳分上壳(epivalve)、下壳(hypovlave),相互嵌套而成。硅藻的外壳布满规则排列、结构复杂的小孔。硅质壳的外形和小孔的排列模式是硅藻的主要分类依据。传统上硅藻根据外壳的对称性可分为中心辐射对称的中心硅藻(Centric diatom)和两侧对称的羽纹硅藻(Pennate diatom);后者根据是否具有壳缝(raphe)又可以分为有壳缝硅藻和无壳缝硅藻。
硅藻是多样性最高的单细胞真核藻类,现存超过250属,估计有10万余种。硅藻细胞大小跨度极大,从小于1微米至几毫米不等。硅藻分布于几乎所有水体环境中,从热带到极地,从强酸性到强碱性,从淡水到盐湖,从两极到温泉,甚至在土壤、树木的表面都有硅藻的踪迹。硅藻最显著的特征是其多样的硅质外壳(frustule)。硅藻的外壳分上壳(epivalve)、下壳(hypovlave),相互嵌套而成。硅藻的外壳布满规则排列、结构复杂的小孔。硅质壳的外形和小孔的排列模式是硅藻的主要分类依据。传统上硅藻根据外壳的对称性可分为中心辐射对称的中心硅藻(Centric diatom)和两侧对称的羽纹硅藻(Pennate diatom);后者根据是否具有壳缝(raphe)又可以分为有壳缝硅藻和无壳缝硅藻。
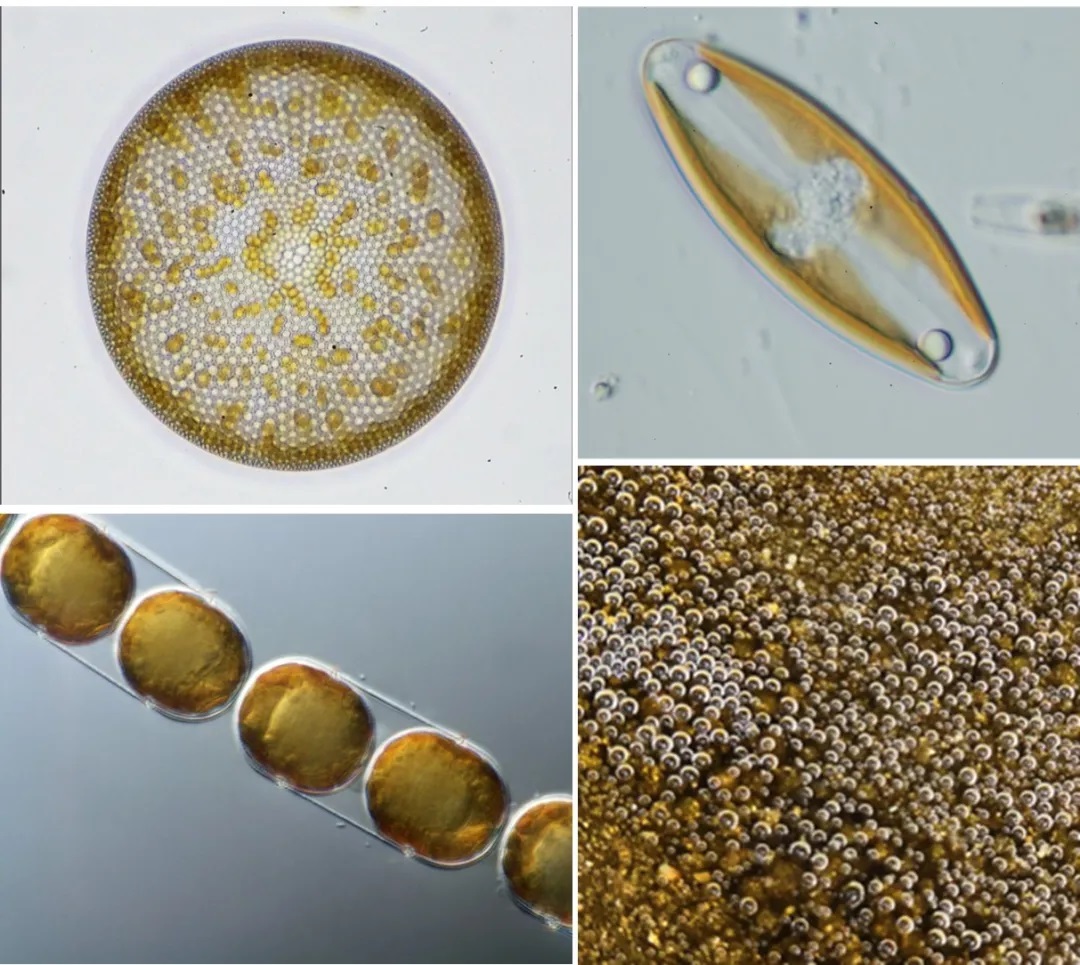
图1 硅藻细胞形态,右下为大量硅藻进行光合作用产氧产生的气泡
会跑的“植物”
根据生境不同,硅藻主要分为浮游硅藻和底栖硅藻。浮游硅藻主要是中心硅藻,而羽纹硅藻大多是底栖类群。浮游硅藻多具有较长的毛或由多个细胞连接形成链状,以增加在水体中漂浮和吸收营养盐的能力。羽纹硅藻则多为底栖类群,其中无壳缝硅藻多通过分泌胞外粘液附着在基质表面,而有壳缝硅藻则具有较强的移动能力,这主要归功于壳缝的出现。硅藻的壳缝如同鸟类的翅膀,使硅藻能够趋利避害,主动适应环境的变化,为硅藻提供了巨大的演化优势,也被认为是底栖硅藻高多样性的主要原因之一。
会跑的“植物”
根据生境不同,硅藻主要分为浮游硅藻和底栖硅藻。浮游硅藻主要是中心硅藻,而羽纹硅藻大多是底栖类群。浮游硅藻多具有较长的毛或由多个细胞连接形成链状,以增加在水体中漂浮和吸收营养盐的能力。羽纹硅藻则多为底栖类群,其中无壳缝硅藻多通过分泌胞外粘液附着在基质表面,而有壳缝硅藻则具有较强的移动能力,这主要归功于壳缝的出现。硅藻的壳缝如同鸟类的翅膀,使硅藻能够趋利避害,主动适应环境的变化,为硅藻提供了巨大的演化优势,也被认为是底栖硅藻高多样性的主要原因之一。
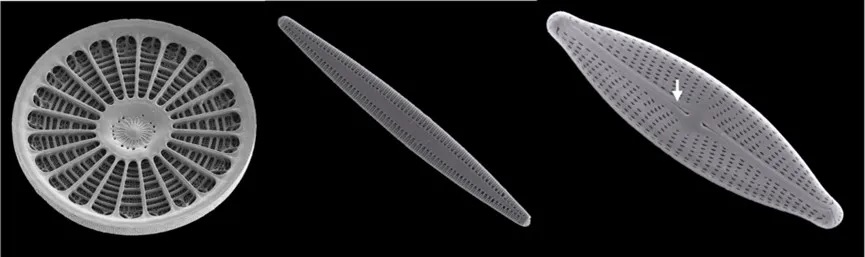
图2 硅藻电镜照片:左中右依次为:中心硅藻、无壳缝羽纹硅藻、有壳缝羽纹硅藻,箭头所示为壳缝

图3 硅藻表面孔纹结构
套娃的循环
硅藻通常进行半保留细胞复制,即在母细胞壳的内部生成新生细胞的外壳,所以随着复制的不断进行,新生细胞的外壳不断变小。当其变为母细胞的大约一半时,达到麦克唐纳-普菲策尔极限(MacDonald-Pfitzer limit),此时会诱发硅藻进行有性生殖。除重组遗传物质外,硅藻有性生殖的另一个重要作用是通过形成复大孢子恢复细胞的原始大小。复大孢子通常由硅质的鳞片和环带组成,在其内部会形成初生细胞,也就是恢复了原始大小的细胞,随后开启细胞分裂增殖、逐渐变小的过程。如果不经过有性生殖,大多数硅藻会逐渐变小直至死亡。然而,也有少数硅藻可以不经有性生殖,直接形成复大孢子恢复细胞的大小。
黄色的海洋
硅藻是现代海洋中多样性最高、最具优势的藻类类群,贡献了约40%的海洋初级生产力和全球20-30%的氧气。硅藻的产氧量甚至高于有“地球之肺”之称的热带雨林。作为海洋生物碳泵的主要组成,海洋中40%的碳,由硅藻通过沉降作用输送至深海,进行长期封存。此外,硅藻在硅、氮的地球化学循环中也占有十分重要的地位,维系着地球生态系统的稳定。
硅藻是藻类中相对较晚演化出现的类群,最早的化石可追溯至白垩纪晚期。此时,海洋中多样性最高的是甲藻和颗石藻。硅藻、颗石藻、甲藻都含有叶绿素a和c,是由二次内共生演化出的藻类类群,它们均使用叶黄素作为天线色素,色素体呈现黄色或褐色。颗石藻和甲藻最早出现于中生代三叠纪,随后多样性逐渐增加,在白垩纪达到顶峰。在白垩纪大灭绝后,步入新生代以来,硅藻的多样性迅速增加,初步取代了甲藻和颗石藻成为海洋中多样性最高的类群。那么是什么原因使硅藻取代甲藻和颗石藻成为新生代海洋的主宰呢?
硅藻最大的特点是具有硅质的坚硬外壳,而甲藻和颗石藻的细胞壁则分别由纤维素和钙质组成。结构复杂的硅质外壳更有利于分散压力,使其可以承受很高外界压力。研究表明,硅藻的外壳可以承受1–7 N/mm2的压强,相当于100–700 t/m2,其机械强度相当于皮质骨和医用牙科材料。在现代海洋中,浮游植物的生物量平均每2–6天就会被捕食者消耗完毕,只有通过不断的分裂产生新的细胞方可维持其生物量。中生代和新生代时期,很可能也存在类似的被捕食率。面对如此高的捕食选择压力。硅藻坚硬的外壳可以降低硅藻被捕食的概率,带来巨大的竞争优势。另外,硅藻分裂增殖过程中,对于恢复硅质外壳大小的需求,使得有性生殖更加必要和频繁,从而带来更多的基因重组机会和演化优势,使得硅藻在物种数量上和物种形成速率上都明显高于其他藻类。
此外,硅藻外壳布满纳米级精细的小孔结构可以更好的利用可见光,并最大限度地减少有害紫外线辐射,降低光损伤。同时,硅藻具有一种特殊的捕光天线蛋白“岩藻黄素-叶绿素a/c蛋白复合体”,使硅藻具有出色的蓝绿光捕获能力和极强的光保护能力,即使在弱光环境中也能生存。另外,硅藻还演化出一种营养盐储藏液泡,可以维持高浓度的氮和磷供给,即使在短期之内没有外源营养盐的情况下,也可以维持细胞分裂。这一结构使硅藻可以在富营养水体或水体混合强烈、营养盐波动剧烈的海域,比其他藻类更具有竞争优势。新生代地质变迁,如德雷克海峡的开放及其后南极环流的形成,导致了季节性营养物质波动,这也为硅藻在海洋浮游植物群落中占据主导地位提供了条件。
硅藻的生存离不开硅元素。海洋中的硅主要来源于大陆岩石的风化,并通过河流输送至海洋。新生代的造山运动如喜马拉雅运动和中新世以来的地壳运动,不仅改变了地球的面貌,同时大大增加了海洋中硅元素的量。此外,在新生代还发生了有趣的草本植物和硅藻的陆海协同演化现象。草本植物比木本植物具有更强的硅积累能力。硅的积累增强了草本植物的竞争力,使其能够在干旱、贫瘠的土壤环境中生存和繁衍。草本植物的根系发达,能够深入土壤,进一步加速硅酸盐矿物的风化。陆地硅酸盐风化的增强导致海洋硅酸盐浓度的增加,促进了海洋中硅藻的繁盛,提高了海洋的初级生产力。草本植物的扩张和硅循环对气候产生了深远影响。硅藻的繁盛有助于将大气中的二氧化碳封存至海洋深处,从而降低大气二氧化碳浓度,对全球气温下降产生积极的作用。
硅藻是现代海洋中多样性最高、最具优势的藻类类群,贡献了约40%的海洋初级生产力和全球20-30%的氧气。硅藻的产氧量甚至高于有“地球之肺”之称的热带雨林。作为海洋生物碳泵的主要组成,海洋中40%的碳,由硅藻通过沉降作用输送至深海,进行长期封存。此外,硅藻在硅、氮的地球化学循环中也占有十分重要的地位,维系着地球生态系统的稳定。
硅藻是藻类中相对较晚演化出现的类群,最早的化石可追溯至白垩纪晚期。此时,海洋中多样性最高的是甲藻和颗石藻。硅藻、颗石藻、甲藻都含有叶绿素a和c,是由二次内共生演化出的藻类类群,它们均使用叶黄素作为天线色素,色素体呈现黄色或褐色。颗石藻和甲藻最早出现于中生代三叠纪,随后多样性逐渐增加,在白垩纪达到顶峰。在白垩纪大灭绝后,步入新生代以来,硅藻的多样性迅速增加,初步取代了甲藻和颗石藻成为海洋中多样性最高的类群。那么是什么原因使硅藻取代甲藻和颗石藻成为新生代海洋的主宰呢?
硅藻最大的特点是具有硅质的坚硬外壳,而甲藻和颗石藻的细胞壁则分别由纤维素和钙质组成。结构复杂的硅质外壳更有利于分散压力,使其可以承受很高外界压力。研究表明,硅藻的外壳可以承受1–7 N/mm2的压强,相当于100–700 t/m2,其机械强度相当于皮质骨和医用牙科材料。在现代海洋中,浮游植物的生物量平均每2–6天就会被捕食者消耗完毕,只有通过不断的分裂产生新的细胞方可维持其生物量。中生代和新生代时期,很可能也存在类似的被捕食率。面对如此高的捕食选择压力。硅藻坚硬的外壳可以降低硅藻被捕食的概率,带来巨大的竞争优势。另外,硅藻分裂增殖过程中,对于恢复硅质外壳大小的需求,使得有性生殖更加必要和频繁,从而带来更多的基因重组机会和演化优势,使得硅藻在物种数量上和物种形成速率上都明显高于其他藻类。
此外,硅藻外壳布满纳米级精细的小孔结构可以更好的利用可见光,并最大限度地减少有害紫外线辐射,降低光损伤。同时,硅藻具有一种特殊的捕光天线蛋白“岩藻黄素-叶绿素a/c蛋白复合体”,使硅藻具有出色的蓝绿光捕获能力和极强的光保护能力,即使在弱光环境中也能生存。另外,硅藻还演化出一种营养盐储藏液泡,可以维持高浓度的氮和磷供给,即使在短期之内没有外源营养盐的情况下,也可以维持细胞分裂。这一结构使硅藻可以在富营养水体或水体混合强烈、营养盐波动剧烈的海域,比其他藻类更具有竞争优势。新生代地质变迁,如德雷克海峡的开放及其后南极环流的形成,导致了季节性营养物质波动,这也为硅藻在海洋浮游植物群落中占据主导地位提供了条件。
硅藻的生存离不开硅元素。海洋中的硅主要来源于大陆岩石的风化,并通过河流输送至海洋。新生代的造山运动如喜马拉雅运动和中新世以来的地壳运动,不仅改变了地球的面貌,同时大大增加了海洋中硅元素的量。此外,在新生代还发生了有趣的草本植物和硅藻的陆海协同演化现象。草本植物比木本植物具有更强的硅积累能力。硅的积累增强了草本植物的竞争力,使其能够在干旱、贫瘠的土壤环境中生存和繁衍。草本植物的根系发达,能够深入土壤,进一步加速硅酸盐矿物的风化。陆地硅酸盐风化的增强导致海洋硅酸盐浓度的增加,促进了海洋中硅藻的繁盛,提高了海洋的初级生产力。草本植物的扩张和硅循环对气候产生了深远影响。硅藻的繁盛有助于将大气中的二氧化碳封存至海洋深处,从而降低大气二氧化碳浓度,对全球气温下降产生积极的作用。
硅藻有什么用?
这大概是最多被问到的问题,也是作为分类研究工作者最为头疼的问题。任何生物经过亿万年的演化,必然有其适应环境的精妙之处。硅藻作为地球上最为成功的生物之一,必然有值得学习利用的地方。硅藻具有精美、坚硬而多孔外壳,在牙科材料、滤材、研磨材料、添加剂等方面的应用已十分广泛,最为人所熟知的是硅藻土。硅藻土是硅藻的外壳沉积形成的硅质生物沉积岩。正是通过添加硅藻土,大幅提高了硝化甘油稳定性和爆炸力,才奠定了诺贝尔“炸药大王”的地位和今天诺贝尔奖的基础。此外,在啤酒酿造过程中,硅藻土也是最常用的过滤解决方案,用于过滤啤酒中的酵母细胞和杂质,使啤酒清澈透明,并可长期保存。
硅藻壳的物理特性在微观纳米材料和生物技术领域同样具有十分广阔的应用前景。例如,中心硅藻的外壳已被成功地用作微透镜,并能够在衍射极限下压缩光;硅藻壳高效收集光的能力可以用于开发新一代仿生太阳能电池;硅藻壳的光致发光可被用于制作光学与生物传感器;功能化的硅藻壳可用作药物递送载体;对硅藻壳的适当修饰可以获得高效的纳米结构半导体器件等等。然而,硅藻最令人着迷的潜力,是通过基因技术调控硅藻壳的形态、几何形状、孔隙分布,以大规模高效合成纳米材料,这也是许多研究人员正在努力的方向。
除硅质壳外,硅藻作为淡水和海洋生态系统中主要的初级生产者,也是水产养殖中重要的天然饵料。硅藻富含EPA、DHA、岩藻黄素与多种高附加值活性物质,在健康食品、生物医药方面也有十分广阔的应用前景。硅藻高效吸收营养盐和光合固碳能力可应用于废水处理和碳封存技术的开发等等。
所有这些应用都是建立在对硅藻生物特性的认知之上。然而,目前我们对硅藻的认知仍十分有限,10万多种硅藻,每一种都有独特的生物学特性,但目前命名的硅藻物种仅有1万余种,仍有90%以上的硅藻等待人类来揭开其神秘的面纱。因此,首先要做的是认识和了解他们,如同每一个婴儿首先要取名字一样,有了名字才能被当做一个独立的个体被对待和认识。硅藻也是如此,发现、描述、命名、认知,这是所有研究和应用的起点和基础,也是分类学的主要工作。相比于陆地动植物,认识藻类的任务才刚刚起步,任重道远。
主要参考文献
Behrenfeld, M. J., Halsey, K. H., Boss, E., Karp‐Boss, L., Milligan, A. J., & Peers, G. (2021). Thoughts on the evolution and ecological niche of diatoms. Ecological Monographs, 91(3), e01457.
De Tommasi, E., Gielis, J., & Rogato, A. (2017). Diatom Frustule Morphogenesis and Function: A Multidisciplinary Survey. Marine Genomics,35, 1–18.
Falkowski, P. G., Katz, M. E., Knoll, A. H., Quigg, A., Raven, J. A., Schofield, O., & Taylor, F. J. R. (2004). The Evolution of Modern Eukaryotic Phytoplankton. Science, 305(5682), 354–360.
Round, F. E., Crawford, R. M., & Mann, D. G. (1990). Diatoms: Biology and morphology of the genera. Cambridge University Press.
井上勲. (2006). 藻類30億年の自然史: 藻類からみる生物進化. 東海大学出版会.
(李宇航 供稿)
作者简介
李宇航,中国科学院海洋研究所海洋生物分类与系统演化实验室副研究员。长期从事硅藻分类、多样性与系统演化研究,发现并描述硅藻新种17种,发表SCI收录论文36篇。主持国家自然科学基金面上项目、青年基金项目等课题。